Leaping Capabilities



Leaping Capabilities |
|
Leaping Speeds
The literature pertaining to leaping capabilities by fish is limited, and often uses observed leap heights to back calculate burst swimming velocities attained prior to leaving the water (Aaserude and Orsborn 1986). The equation used to back calculate leap height is usually some form of the simple projectile equation:
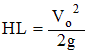
where,
HL = leap height
Vo = initial velocity of the fish
g = gravitational acceleration
Aaserude and Orsborn (1996) working with pacific salmonids used burst velocities of short duration to predict leap height and found that they closely matched the range of observed leap heights. Stuart (1962) and Reiser and Peacock (1985) have also stated that leaps use burst speeds of short duration. FishXing uses short duration burst velocities (5 sec burst speeds) to estimate the speed at which fish leave the water when leaping. In FishXing the Maximum Leap Speed is maximum velocity at which a fish exits the water when leaping at the culvert outlet. See the Leaping Calculations section to view the calculations.
Factors Affecting Leaping Success
Besides swimming speed and fall height several other factors appear to affect leaping success, including water velocity, turbulence in the take-off pool, the depth of the take-off pool, the ratio of pool depth to fall height, and where the leap is initiated.
Flow rate may affect the motivation of fish to leap at outfalls and when it does there appears to differences between species and life history stages. Stuart (1962) studying small fish (minnows, and juvenile salmon: 6-15 cm) and Lauritzen (2002) studying kokanee salmon found that passage attempts increased with flow until the flow rate exceeded the swimming capacity of the fish. Conversely, Powers and Orsborn 1985 observed that chum and coho could sometime be stimulated to move upstream by increasing the flow, but they also leaped over outfalls when the flow was reduced to just a trickle.
It seems that different species of fish and life stages may use different leaping techniques, which may lead to differing physical conditions needed to support leaping. Stuart (1962) found that minnows and juvenile salmon initiated their leaps from the surface of the water following a quick body and tail flexion that bent their bodies into a C-shape. These fish also always leaped from the standing wave created by the upwelling of water just downstream from where the falling water stuck the pool surface. On the other hand, Lauritzen (2002) found that adult kokanee salmon initiated their leaps from the bottom of the standing wave with an S-shaped curve of their bodies followed by burst swimming to the surface. These fish also mainly leaped from the standing wave. Aaserude and Orsborn (1986) found that smaller resident trout initiated their leap from the standing wave more frequently than did adult coho or chum salmon. These authors suggest that this difference may be because the upwelling presented a more influential flow condition for trout due to their smaller mass. The depth that is needed in the outfall pool is likely to be related to the leaping technique used.
The turbulence in the outlet pool will influence the fish's ability to jump. As the fish leaps it pushes against the water with its tail. In turbulent water air is entrained in the water column, and instead of the fish’s tail pushing against water it pushes against air. Because air has a much lower density the fish’s tail produces less thrust. In addition, fish seem to use the flow to help direct their leaps, when the water is turbulent their leaping is less directed. Turbulence in the outlet pool is a function of factors such as water velocity, and outlet pool depth in relation to the drop height.
![]() FishXing Version 3.0 Beta, 2006
FishXing Version 3.0 Beta, 2006