Fish Swimming and Swim Speed Tests



Fish Swimming and Swim Speed Tests |
|
Fish Swimming performance is one of the basic FishXing model inputs and it is important to understand how swim speed estimates were developed and how accurate they might be. Most of the swim data available in the literature is derived from laboratory experiments under controlled and usually non-volitional swimming conditions.
While a thorough discussion of this topic is beyond the scope of this Help manual, the following is a brief outline of the issues involved in estimating swim speeds. More in depth discussion can be found in reviews of swimming speed tests by Beamish (1978), Cech (1990), and Hammer (1995).
Test Chambers
Beamish (1978) describes various test procedures and discusses conditions that produce variable results. Swim speed tests are usually carried out in test chambers of two basic types: in one, the chamber rotates, and in the other, water flows through a stationary chamber.
The test chamber can affect the estimate of the water velocity experienced by the fish. Rotating chambers create complex flow patterns that can affect estimates of water velocity. Where the water moves and the chamber stays stationary, the chamber is often a tube (small water volume) or flume (large water volume). Unless flow straighteners are used flow through the tube or flume can also be complex and affect the estimate of water velocity. Even when laminar flow is achieved, precise description of swimming capacity is difficult because not only does the precise location of the fish need to be monitored, but also the velocity that can be assigned to each location (Beamish 1978).
Fish Size
Another consideration when using test chambers to estimate swim speed, is the size of the fish relative to the size of the test chamber. When the fish’s body occupies a significant portion of the chamber, the chamber effectively becomes narrower resulting in an accelerated flow over the body of the fish. Additionally, fish experience a higher than expected drag in an enclosed flume as compared to freestream velocity. The additional drag comes from horizontal buoyancy and solid blocking effects (Beamish 1978). Corrections for these effects have been described by Webb (1971).
Test Procedures
The test procedure can also affect the estimates of swimming speeds. For example, the length of time fish are allowed to recover from handling stress after being transferred to the test chamber has been shown to affect swimming speed. The rate and magnitude of water velocity increases also can influence test results. Hammer (1995) provides a good summary of this topic.
There are three basic types of test procedures:
1. Fixed velocity or endurance tests,
2. Increasing velocity or critical velocity tests, and
3. Volitional swimming tests that involve measuring the distance traveled against a water velocity in an open channel.
Endurance Tests
In the fixed velocity or endurance test, velocity is abruptly or gradually stepped up to the prescribed level. After the test velocity is reached, the time to exhaustion is measured. When 50 percent of the fish have fatigued, the mean maximum sustainable speed is reached (Brett 1967).
Critical Velocity Tests
In increasing velocity or critical velocity tests, the fish is subjected to increasing velocities in a series of steps (velocity increments) with each step being maintained for a specific period of time (time between increments). Brett (1964) calculated the fatigue speed or critical velocity (Ucrit) as:
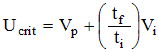
where,
Vi = the velocity step (cm/s)
Vp = the penultimate velocity reached at fatigue (cm/s)
tf = the time lapsed from the velocity increase to fatigue (s)
ti = the time between velocity increments (s)
Hammer (1995) provides a literature review on the effect of velocity increment and the time between increments. Farlinger and Beamish (1977) demonstrated that changes in both velocity increment and time between increments have a marked effect on the swimming speed of largemouth bass (Micropterus salmoides). Hammer (1995) concludes from his review of the literature that velocity intervals shorter than 15 to 20 minutes have a marked effect on the estimate of critical velocity. However, he finds no reason for the time to be as long as the 60 minutes Brett (1964) recommended, and states that 30 minutes intervals seem to be appropriate.
It appears that following a velocity increase fish exhibit a period of restlessness and Webb (1971, in Hammer 1995) suggest that some anaerobic energy production occurs after each increase. He concludes that the time required to adjust to the higher speeds with time increments less than 20 minutes may take up a significant portion of the time at that increment and affect the results of the test. Webb (1971) further suggests that fixed velocity and increasing velocity tests measure different forms of exhaustion and should not be compared.
Volitional Swimming Tests
The last form of test involves fish swimming a measured distance against a velocity challenge. Weaver (1963, 1965) tested the ability of migrating anadromous salmonids to ascend an experimental fishway. Recently, Castro-Santos (200?) and Peake (2004) questioned the validity of swim speed estimates derived from data collected using fixed or increasing velocity tests where fish are forced to swim in enclosed tubes at prolonged speeds. Other authors have noted that fish do not swim steadily at prolonged speeds as they are forced to do in most swim speed tests. Routine swimming by fish is often unsteady. At low speeds, “stroke-and-glide” (Swanson et al. 1998), and at high speeds, “burst-and-coast” swimming behavior (Hitch and Bratty 2000) has been observed in fish. These behaviors are believed to increase endurance and reduce energy expenditure (Webb 1994 and Weihs 1974).
Some fish may not be suited to swimming steadily at prolonged speeds. Swanson et al. (1998) measured the critical swimming velocity and behavioral a kinematic limitation on swimming at submaximal velocities in delta smelt (Hypomesus transpacificus). They found that delta smelt exhibited swimming failures at velocities that corresponded to the transition between ‘stroke and glide’ swimming and continuous swimming. Delta smelt were unable or unwilling to swim steadily in the flume within the transition velocity range. They speculate that delta smelt may not swim at these speeds under natural conditions.
Peake (2004) argued that more realistic swimming speeds are generated by testing fish in a 50-meter raceway than in a respirometer tube using increasing velocity tests. He found that fish increased their swimming speed as the water velocity increased. The swimming speeds he found in the raceway were almost twice those predicted from the critical velocity tests. He speculated about the causes of the estimate differences including decreased tail beat amplitude due to confinement in the respirometer tube, but concluded that the explanations presented did not seem satisfactory and were likely caused by a combination of behavioral, physiological, and hydraulic conditions.
There is some evidence that migratory teleosts swim at a constant ground speed through a wide range of water velocities spanning both prolonged and burst swimming speeds. Swimming at constant ground speed may optimize the distance traveled in some, but not all cases (Castro-Santos 2002).
FishXing and Swim Speeds
Rather than a constant ground speed, FishXing assumes a constant swimming speed, either at their estimated prolonged or burst swimming speeds based on the water velocity. Both the validity and effect of this assumption is the subject of current study, and we expect that estimates of swimming speeds will improve over time. Most of the constant and calculated swim speed estimates in FishXing’s literature Swim Speeds Table were derived from fixed velocity or increasing velocity tests. The limitations of these tests in accurately predicting swimming speed should be understood before using this model.
Variability Between Individuals and Populations
The biological parameters and criteria used in an analysis are typically intended to represent the overall population of a specific species and lifestage. However, several researchers have commented on the wide range in swimming speeds that individuals display (Berry and Pimentel 1985, Kovacs and Leduc 1982, Mcdonald et al., 1998, and McNeish and Hatch 1978) even when the major factors affecting speeds such as species, size, life stage and stock are taken into account. McDonald et al. (1998) suggested that the individual differences were probably due to real inter-individual physiological differences. Berryand Pimentel (1985) suggest that the individual differences are probably due to different levels of motivation and stress. Because swimming speeds are variable, the species and life stage mean swimming speed may not be the best estimated to use to ensure that most of the individuals within a species and life stage can pass the culvert. It is important to consider this variability when choosing the swimming speed for the species and life stage that you will be analyzing with FishXing.
See: Time to Exhaustion, Swim Categories, Swim Speeds Table
![]() FishXing Version 3.0 Beta, 2006
FishXing Version 3.0 Beta, 2006